Duplicación cromosómica y el origen de nueva información biológica: Estudio de caso sobre las globinas
21 octubre 2011 — Aquellos que hemos estado involucrados en la discusión y debate acerca de DI/evolucionismo desde hace cierto tiempo estaremos bien familiarizados con el modelo neodarwinista más de moda para el origen de una nueva información biológica: la duplicación cromosómica y la consiguiente divergencia. Las duplicaciones de genes suceden generalmente debido a un «sobrecruzamiento defectuoso», una especie de sobrecruzamiento que ocurre durante la meiosis entre cromosomas homólogos mal alineados. Este proceso tiene como resultado la deleción de una secuencia en una hebra, y su sustitución con una duplicación procedente de su cromosoma homólogo. El modelo de duplicación cromosómica y divergencia mantiene esencialmente que, después de una duplicación de un gen, en tanto que una copia del mismo retiene su función original, la otra copia queda liberada del freno selectivo, y queda por ello libre para mutar a un ritmo más rápido y para explorar el espacio de secuencias en busca de alguna nueva función.
Recientemente, un amigo me pidió datos acerca de este tema tan interesante e importante, y esto me ha llevado a escribir un breve artículo en el que recojo algunos de mis pensamientos sobre este tema.
Recientemente, un amigo me pidió datos acerca de este tema tan interesante e importante, y esto me ha llevado a escribir un breve artículo en el que recojo algunos de mis pensamientos sobre este tema.

El propósito de la hemoglobina en la sangre es transportar oxígeno desde los órganos de respiración a los tejidos del cuerpo, donde se libera el oxígeno para proporcionar energía para el organismo. La hemoglobina sirve también para recoger el dióxido de carbono y devolverlo a los pulmones (o branquias en el caso de los peces) de modo que pueda ser expulsado del organismo. Cuatro cadenas polipeptídicas componen el tetrámero funcional de la hemoglobina. Dos de ellas son idénticas en estructura (y se designan como la cadena α), y las otras dos son también idénticas entre sí (y se designan como una cadena β). En el diagrama de la estructura de la hemoglobina (más arriba), el rojo designa las subunidades α, y el azul las subunidades β. Los grupos hemo, que contienen hierro, se representan en verde. La mioglobina (específica de las células musculares) posee una afinidad aun mayor para el oxígeno que la hemoglobina. La mioglobina actúa almacenando el oxígeno, y lo retiene en los músculos cardíaco y esqueléticos.
Ahora bien, quiero exponer que hay al menos cinco dificultades que se levantan contra una evolución de las globinas mediante duplicación cromosómica y divergencia:
- La cuestión de la función biológica de los intermedios que se proponen.
- Cambios complementarios que involucrarían a la regulación de la expresión génica.
- Limitación temporal asociada al hallazgo de una función susceptible de selección para el gen duplicado.
- El problema de la fragilidad.
- Problemas de convergencia.
Pasemos a examinar cada uno de estos puntos.
La función biológica de los intermedios que se proponen
La función biológica de los intermedios que se proponen
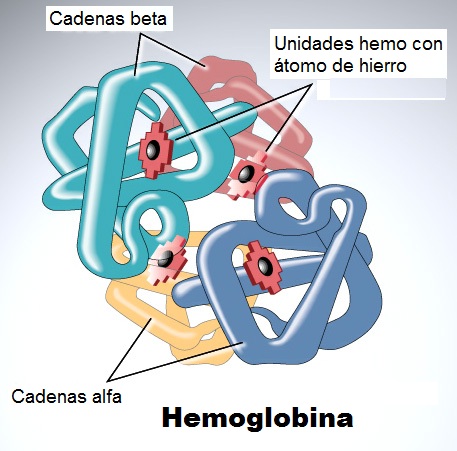
Pasemos a considerar la divergencia entre las cadenas α y β de la hemoglobina. Después de la duplicación del gen inicial de la hemoglobina, cada copia tiene que divergir simultáneamente, y de formas complementarias a fin de asegurar un tetrámero funcional. Por una parte, una característica de la hemoglobina es que ambas cadenas poseen aminoácidos hidrófobos esenciales para la asociación de las subunidades (en marcado contraste con la mioglobina que, al ser una proteína soluble en agua, posee mayormente aminoácidos hidrófilos en el exterior de su estructura plegada). Existe la extendida opinión de que la primitiva hemoglobina pudo haber sido un monómero parecido a la mioglobina, en posesión de aminoácidos externos hidrófilos, algunos de los cuales hubieran sido posteriormente sustituidos por aminoácidos hidrófobos cuando evolucionaron las dos diferentes cadenas. Por ejemplo, en su libro Hemoglobin: Structure, function and evolution [La hemoglobina: Su estructura, función y evolución] (1983), Richard Dickerson e Irving Geis arguyen que algunas de estas sustituciones tuvieron lugar antes de la duplicación cromosómica: es decir, que la primitiva hemoglobina evolucionó para formar un tetrámero (o quizá un dímero) antes de divergir a las cadenas α y β. Pero una sugerencia así es dudosa a la luz del hecho de que uno podría esperar ver estos aminoácidos hidrófobos en posiciones similares si aparecieron antes de la divergencia de las hemoglobinas. Pero los aminoácidos hidrófobos en el exterior de los polipéptidos son diferentes y aparecen situados en diferentes posiciones en las cadenas α y β. Esto es lo que promueve su complementariedad: La condición hidrófoba no se necesita en el mismo lugar para cada polipéptido respectivo, sino sólo en aquellos sitios que establecen contacto con las otras cadenas. Las cadenas α no pueden hacer contacto entre sí, a diferencia de las cadenas β que sí que lo establecen. Cuando no se producen las cadenas β (como sucede en la β-talasemia, un trastorno que resulta a menudo fatal), las cadenas α no asociadas generalmente se degradan. Pero en el caso de la α-talasemia, donde son las cadenas α las que están ausentes, las cadenas β sí que se asocian. Esto presenta una dificultad diferente, porque éstas ligan el oxígeno con una afinidad similar a la de la mioglobina. Esto significa que el oxígeno no será liberado a los tejidos —una vez más con consecuencias fatales.
Dada la plétora de trastornos patológicos que se pueden atribuir a un cambio en sólo un aminoácido en estas cadenas (con lo que se trastoca la cooperación entre las cadenas), debería resultar bien evidente que muchos aminoácidos en ambas cadenas son esenciales para el funcionamiento correcto de la hemoglobina tetrámera. Además, muchos de los aminoácidos implicados en esta interacción están situados en diferentes posiciones en las secuencias polipeptídicas (o bien se encuentran aminoácidos diferentes en posiciones análogas).
Así, espero haber demostrado en lo referente a nuestro actual caso bajo estudio (esto es, las globinas), que para poder considerar creíbles los árboles genealógicos genéticos, es necesario demostrar que hay una probabilidad suficientemente elevada de que todas las secuencias ancestrales confieran alguna especie de ventaja selectiva. Pero lo que se ha demostrado una y otra vez es que pasar de un conjunto de aminoácidos conservados a otro —como es necesario para la producción mediante divergencia de proteínas con diferentes funciones— constituye un salto demasiado grande a través del espacio de secuencias.
Como queda documentado por Axe y Gauger (2011), incluso un cambio aparentemente trivial de la función Kbl a BioF exige al menos siete mutaciones coordinadas, lo que sitúa la transición bien fuera del alcance de un proceso darwinista dentro del tiempo que se asigna a la edad de la tierra. El artículo de Axe y Gauger estudia la superfamilia de transferasas PLP-dependientes. Han identificado un par dentro de la superfamilia con una estrecha similitud estructural pero sin función superpuesta. Las enzimas escogidas fueron Kbl (involucrada en el metabolismo de la treonina) y la BioF (que forma parte de la ruta biosintética de la biotina). Usaron un proceso en tres etapas para identificar las secuencias que con más probabilidad conferirían un cambio de función.
Este proceso en tres etapas involucraba:
- El uso de comparaciones estructurales y de secuencias de las dos enzimas para identificar aminoácidos candidatos con mayor probabilidad de ser funcionalmente significativos.
- La mutación de estos aminoácidos en la BioF, haciéndolos parecidos a Kbl, y comprobación de la pérdida de la actividad de BioF.
- El ensayo de si el cambio de estos grupos en Kbl para que se asemejen a BioF capacitará a la Kbl para que sustituya la función de la BioF.
De este modo estimaron que se necesitarían siete o más mutaciones para convertir la Kbl a la función de la BioF.
El artículo de Axe y Gauger no es un resultado aislado. Una reseña en Natureinformaba de que cambiar la actividad química de una enzima puede necesitar múltiples mutaciones neutrales o deletéreas.
El intercambio de reacciones catalizadas por miembros de superfamilias mecanísticamente diversas se podría considerar como ejercicios «fáciles» en (re)diseño: si la Naturaleza lo hizo, ¿por qué no podremos nosotros? ... Como anécdota, desde entonces se han realizado muchos intentos de intercambio de actividades en superfamilias mecanísticamente diversas, pero se han conseguido pocos éxitos.
Cuando estos resultados se toman en cuenta en el contexto de las predicciones de genética de poblaciones con respecto al tiempo de espera para múltiples mutaciones coordinadas no adaptativas necesarias para facilitar una determinada transición (p. ej. véase Axe 2010), la situación en que se encuentra el neodarwinismo parece insostenible.
Muchos lectores serán conocedores del famoso artículo en JMB de 2004 de Douglas Axe sobre la β-Lactamasa. Axe emprendió su investigación con la intención de determinar el predominio de las variantes de secuencia con una concreta firma hidropática que pudiera formar una estructura funcional de entre el espacio de posibilidades combinatorias. Axe comenzó con una variante sumamente débil (sensible a la temperatura), implicando que sería de esperar que un nuevo pliegue en evolución sería deficientemente funcional. Con ello, Axe buscaba detectar variantes operando en el nivel inferior —el umbral, por así decirlo— de la detectabilidad. Axe buscaba proporcionar una estimación de la escasez de los pliegues funcionales en todo el espacio de secuencias, que él da como de 1 en 10^77. Esta estimación se extrapoló a partir de la cantidad de variantes que podían realizar la función, por débilmente que fuese, de la enzima TEM-1 β-Lactamasa.
La investigación de Axe no constituye tampoco un resultado aislado. Por ejemplo,un estudio, publicado en Nature en 2001 por Keefe y Szostak, documentaba que se precisaba de más de un millón de millones de secuencias aleatorias para poder tropezar con una proteína de tamaño modesto captadora de ATP. Además,Taylor et al., en su artículo de 2001 en PNAS, obtienen un resultado similar. Dicho artículo examinaba la corismato mutasa de tipo aroQ, y llegaba a un predominio similarmente bajo (donde se daba un valor de 1 en 10^24 para la enzima de 93 aminoácidos, pero, al ajustar para reflejar un residuo de la misma longitud que la sección de 150 aminoácidos analizada de la β-lactamasa, el resultado se da como 1 en 10^53). Otro artículo de Sauer y Reidhaar-Olson (1990) se refería a «el alto nivel de degeneración en la información que especifica un determinado pliegue de proteína», que él da como de 1 en 10^63. También recomiendo a los lectores a examinar el excelente artículo de reseña de Douglas Axe en Bio-complexity donde trata acerca de este tema de forma más detallada, así como la lectura de la obra recién publicada The Nature of Nature — Examining The Role of Naturalism in Science, que es sumamente accesible al no especialista. Axe también ha publicado una entrada de blog en el sitio web de Biologic Institute, donde aborda algunas críticas de su artículo de 2004 en JMB.
En su artículo de 2010, Douglas Axe argumenta (sobre la base de un modelo de genética de poblaciones de bacterias) que la duplicación cromosómica y reclutamiento, como modelo para la evolución de nuevos genes, es algo muy limitado. Funciona sólo si se precisa de muy pocos cambios para llegar a una nueva función susceptible de selección. Si el gen duplicado tiene un coste de aptitud ligeramente negativo, la máxima cantidad de mutaciones (además de la duplicación misma) que puede necesitar una nueva innovación en una población bacteriana es de dos o menos. Si la duplicación es sin coste, el número de mutaciones pasa a seis o menos.
Cambios complementarios que involucrarían la regulación de la expresión génica
A menudo parece que los científicos están tan interesados en la similitud de estructura y función de la hemoglobina y de la mioglobina que descuidan completamente la realidad de que se producen en tejidos completamente diferentes: la médula ósea y los músculos, respectivamente. La función de la mioglobina es posibilitada por su más elevada afinidad por el oxígeno. Las modificaciones de la secuencia de aminoácidos de la proteína, para poder ser convertida de hemoglobina a mioglobina, hubieran tenido que ir acompañadas de cambios complementarios en las secuencias reguladoras con el fin de asegurar la producción de la mioglobina en el músculo, donde se necesitaba, en lugar de en la médula ósea donde se producen los glóbulos rojos de la sangre. La mioglobina presente en los glóbulos rojos, o eritrocitos, no proporcionaría ninguna ventaja selectiva. En realidad, sería dañina para el organismo debido a que se uniría demasiado fuertemente con el oxígeno y no lo liberaría a los tejidos (como sucede en el caso de la α-talasemia que se ha descrito más arriba). Este es uno de los puntos más olvidados en los razonamientos acerca de duplicación cromosómica y árboles genealógicos. Otro ejemplo de esto lo tenemos en la pretendida duplicación de la cadena β para dar origen a la versión fetal γ que posee una afinidad mayor para el oxígeno. Un escenario así, naturalmente, tendría que suceder de forma concertada con cambios complementarios en control génico como el que se expresa durante el embarazo, pero la síntesis de la cadena β queda impedida hasta el nacimiento.
Limitación temporal
Hay la extendida creencia de que un gen duplicado no tiene asociados ningún coste o ventaja en el fenotipo: es decir, que selectivamente es neutral. En tal estado, se piensa que el gen es libre de mutar, independiente de limitaciones o presiones selectivas. Cuando un gen previamente codificante de proteína sufre mutaciones deletéreas de modo que ya no codifica un polipéptido útil, dicho gen se convierte en un «pseudogén».
Sin embargo, Kuo y Ochman (2010) presentan un potente desafío a este punto de vista en un artículo publicado en PLoS Genetics, titulado «The Extinction Dynamics of Bacterial Pseudogenes [La dinámica de la extinción de los pseudogenes bacterianos]». Los investigadores examinan los genomas de la especie Salmonella, argumentando que la evolución de los pseudogenes bacterianos no es un proceso totalmente neutral. Al contrario, los pseudogenes bacterianos son a menudo degradados activamente por las fuerzas selectivas. La degradación de genes que no codifican proteínas ni realizan funciones reguladoras, se debe muy probablemente a los costes energéticos de su transcripción a ARNm.
La conclusión de los investigadores es que «Debido a que todos los grupos bacterianos, así como los de las arqueobacterias que se han examinado, exhiben un patrón mutacional sesgado en favor de deleciones y que sus genomas haploides serían más susceptibles a efectos negativos dominantes que impartirían los pseudogenes, es probable que el proceso de eliminación adaptativa de los pseudogenes sea ubicuo entre los procariontes». También sugieren que el principio de la evolución reductora (de recorte de costes) podría extenderse más allá del ámbito de los procariontes, basándose en evidencia de selección sobre el tamaño de los intrones en algunos genomas eucariotas (se supone que también debido al coste energético de la transcripción).
Muchos lectores podrán recordar la publicación de un artículo publicado el año pasado en la revista bio-complexity por Gauger et al. En aquel artículo se llegaba a una conclusión muy parecida. Gauger et al demostraban que el proceso de evolución reproductiva era suficiente para impedir que líneas de E. coli adoptasen una simple ruta en dos etapas a una nueva función de aptitud. En dicho estudio, se «partió» un gen trpA de manera que podía recuperar la capacidad de sintetizar el aminoácido triptófano, revirtiendo sólo dos mutaciones puntuales singulares. Los autores aquí también concluían que el coste de transcribir un gen roto incurre en un coste significativo para la aptitud, y que por ello su eliminación resulta facilitada por la selección positiva.
Entonces, ¿a qué conclusión podemos llegar? Evidentemente, todo ello respalda el concepto —que ha sido expuesto por proponentes del DI durante algo de tiempo— de que existe una significativa limitación en la cantidad de mutaciones que se puede acumular mediante un «paseo aleatorio» antes de la degradación y la pérdida de un gen «roto», lo que limita significativamente la capacidad del «motor de búsqueda» mutacional de explorar y ensayar el vasto océano de posibilidades combinatorias. También sirve como potente recordatorio de la insuficiencia causal del mecanismo darwinista de mutaciones aleatorias y selección natural para explicar los orígenes de dominios y funciones estructurales de proteínas fundamentalmente nuevas.
De hecho, como lo expone Kimura, en el capítulo 10 de su célebre libro The Neutral Theory of Molecular Evolution [La teoría neutral de evolución molecular] (1983):
El proceso que facilita la producción de nuevos genes causará a la vez la degeneración de una de las copias duplicadas. De hecho, la probabilidad de que la duplicación cromosómica lleve a degeneración tiene que ser mucho más elevada que la de llevar a la producción de un nuevo gen que tenga alguna función útil.
El problema de la fragilidad

Las globinas poseen un grupo hemo (véase imagen superior) que contiene hierro. El hierro existe en dos formas oxidadas, es decir, Fe2+ y Fe3+(las formas ferrosa y férrica, respectivamente). Cuando hay oxígeno disponible, el hierro se oxida fácilmente a férrico, Fe3+. Pero, cosa crucial para su función fisiológica, el hierro presente en la hemoglobina y en la mioglobina existe en el estado de oxidación ferroso. En el capítulo 2 de su libro al que he aludido previamente (Hemoglobin: Structure, function and evolution [La hemoglobina: Su estructura, función y evolución]), Dickerson y Geis observan que «El propósito del hemo y de la cadena polipeptídica a su alrededor es impedir la oxidación del hierro ferroso (la metamioglobina, o ferrimioglobina, con un hierro férrico, no liga oxígeno), y proporcionar un hueco en el que el oxígeno pueda caber».
La oxidación del hierro ferroso a la forma férrica, en la hemoglobina y la mioglobina, queda impedida de manera activa por los grupos químicos específicos de los residuos aminoácidos alrededor. Dickerson y Geis describen la fragilidad de este sistema con estas palabras:
Las formas meta [oxidadas] resultaron las más fáciles de obtener por métodos experimentales, y los estados desoxi también se pudieron cristalizar y estudiar con unos métodos experimentales cuidadosos. Las formas oxi resultaron ser más intratables: a no ser que se trate con un cuidado extremo, el O2 oxida el hierro del hemo de Fe2+ a Fe3+ en lugar de simplemente ligarse al mismo, lo que resulta en la forma indeseable de la molécula.
Como sería de esperar, los aminoácidos que rodean el grupo hemo están evolutivamente muy conservados. Además, se suscitan muchos cuadros patológicos como resultado del cambio de un solo aminoácido, lo que resulta en la consiguiente incapacidad del polipéptido para retener correctamente el grupo hemo, permitiéndose así la oxidación del hierro. En muchos casos, el cambio de un sólo aminoácido hace que se altere el posicionado de los aminoácidos contiguos al grupo hemo, de modo que ya no pueden protegerlo de oxidación. ¡Esto significa que los aminoácidos alterados no tienen siquiera que estar en una proximidad especial al grupo hemo!
Problemas de convergencia
Una cosa es contemplar la gigantesca dificultad de la evolución de la globina, pero el asunto se manifiesta aún más intratable cuando se considera que la duplicación para conseguir mioglobinas y hemoglobinas tiene que haber ocurrido al menos dos veces en los agnatos (peces sin mandíbulas) y en los vertebrados mandibulados. Desde época temprana, se concluyó (basándose en comparaciones de secuencias aminoácidas) que las globinas más primitiva eran monoméricas, y que esto llevó a las hemoglobinas predominantemente monoméricas de las diversas líneas de invertebrados (p. ej., insectos, moluscos, peces sin mandíbulas) y que sólo después de esto el gen de la globina dio origen a las separadas mioglobina y hemoglobina de los vertebrados mandibulados. ¡Pero la presencia de la mioglobina se ha documentado en la lamprea, un pez sin mandíbulas (Romero-Herrera, 1979)!
De hecho, hay diversos ejemplos como este. Otro ejemplo es la pretendida divergencia resultante de la duplicación de las cadenas α y β para dar origen a las cadenas de expresión embrionaria ζ y ε. En las aves se pueden encontrar cadenas de hemoglobina similares de expresión embrionaria llamadas π y ρ. Está extendida la creencia de que la fecha de la divergencia es extremadamente remota, y que precedió a la divergencia de los linajes que emprendieron las respectivas rutas hacia los mamíferos y hacia las aves.
En tanto que las cadenas ζ y ε, aunque similares entre sí, son notablemente diferentes de las cadenas α, las modernas cadenas ε y ρ exhiben unos niveles muy superiores de similitud a las cadenas β de mamíferos y aves, respectivamente. Así, se puede concluir que según la tesis evolucionista la divergencia para dar origen a estas versiones embrionarias ha tenido que ocurrir independientemente al menos en dos ocasiones: en la línea de los mamíferos y en la de las aves.
Conclusión
Como resumen, hemos visto que el ámbito para la evolución de genes novedosos y de nuevas proteínas mediante duplicación cromosómica y la consiguiente divergencia o reclutamiento es muy limitado, incluso para facilitar innovaciones funcionales relativamente triviales. Dadas las variedades extremadamente diversas de conformaciones proteínicas que aparecen en los sistemas vivos, la probabilidad de una relación de descendencia de genes —incluso dentro de familias genéticas— puede tratarse con desconfianza y un sano escepticismo. Parece más bien irónico que entre los biólogos haya una buena disposición a aceptar un argumento estadístico en contra de que dos o más proteínas con secuencias similares hayan surgido de manera independiente por azar, pero que se esté completamente en contra de considerar argumentos estadísticos en contra de que puedan haber aparecido por azar en absoluto.
Fuente: Uncommon Descent – Gene Duplication and the Origin of Novel Biological Information: A Case Study of the Globins
Redacción: © Jonathan M - www.uncommondescent.com
Traducción y adaptación: Santiago Escuain — © SEDIN 2011 - www.sedin.org
Usado con permiso del traductor para: www.culturacristiana.org


Comentarios
Publicar un comentario